


摘要
砷是一种广泛存在于自然环境中毒性较强的类金属元素,农田生态系统中的植物(尤其水稻)很容易吸收积累土壤环境中的砷。植物中的砷沿食物链向高等动物传递,威胁人类健康。除 土壤本身的理化性质外,土壤中砷的生物转化也强烈影响砷的生物有效性。目前研究发现异化砷酸盐(As(V))呼吸性还原、细胞质As(V)还原、亚砷酸盐(As(III))氧化、As(III)甲基化和有机砷的去甲 基化在土壤砷的生物地球化学过程中起重要作用。随着分析化学和分子生物学技术的进步,最新研究 发现土壤生物也参与了砷糖、砷糖磷脂、砷甜菜碱、砷代草丁膦、硫代砷等有机砷的合成,其中三价 一甲基砷和砷代草丁膦可作为新型抗生素,但其合成机制及生态学功能有待进一步研究。本文还详细 介绍了为适应复合污染环境微生物通过自身的进化对抗生素和重金属形成的四种共选择抗性机制:共 抗性,交叉抗性、共调控和生物膜感应,特别提出了土壤中砷污染与抗生素抗性相关联这一新的研究 方向。最后对砷生物转化和砷与抗生素共抗机制的未来研究方向做了展望。
关键词:砷氧化还原;砷甲基化;砷其他有机化;抗生素抗性基因
类金属元素砷( As),位于元素周期表第四 周期第 VA 主族,是一种广泛分布的有毒环境物 质,已被美国环境保护署( USEPA)确定为人类 的致癌物质。岩(矿)石风化、火山活动、矿山开 采、冶炼、石油燃烧以及杀虫剂使用等自然或人为 活动将砷释放进入大气、土壤和水体后,经物理、 化学及生物等反应在地球圈层之间及其圈层内部进 行循环[1]。土壤中的砷可经地表径流或渗漏进入 水体,或随着粉尘和烟雾等进入大气在空气中流通 扩散,或通过植物的吸收积累进入食物链影响人体 健康。
植物对砷的吸收效率主要取决于土壤砷的存 在形态及迁移转化。土壤和水环境中的砷主要是无 机态,在有氧环境中砷以砷酸盐(As(V))的形式被吸附于铁铝等氧化物表面而不易移动,难以被 植物吸收[2];在水稻土这种厌氧环境中,砷主要 以亚砷酸盐(As(III))形式存在,As(III)不 易被吸附,流动性较大,很容易进入植物体内,所 以与其他旱作农作物相比, 水稻籽粒中积累了更 多的砷[3]。除了土壤理化性质(如土壤通气性、 pH、氧化还原电位、土壤胶体特性及吸附性等) 外,土壤中的动物及微生物也通过对砷的吸收转化 极大地影响着砷在环境中的迁移转化、形态毒性及 生物有效性[4-5]。土壤微生物对砷的转化作用主要 体现在:对As(V)的呼吸还原和细胞质还原,产 生移动性更强的As(III);对砷的氧化反应,增 强砷与土壤矿物的吸附性进而影响砷的结合态和生 物有效性;对砷的甲基化后挥发土壤砷而降低植物体内所积累砷的生物毒性。此外砷作为毒性较强的 类金属元素与抗生素共存时将增强抗生素抗性基因(antibiotic resistance genes,ARGs)传播扩散及 其出现频率,某些有机砷如MAs( III)和砷代草 丁膦还可作为新型抗生素。本文主要介绍土壤砷的氧化/还原、甲基化/去甲基化以及砷其他有机化过 程的最新研究进展(图1),和四种微生物共选择 抗性机制:共抗性、交叉抗性、共调控和生物膜感 应,并提出了土壤中砷污染与抗生素抗性相耦合这 一新的研究方向。
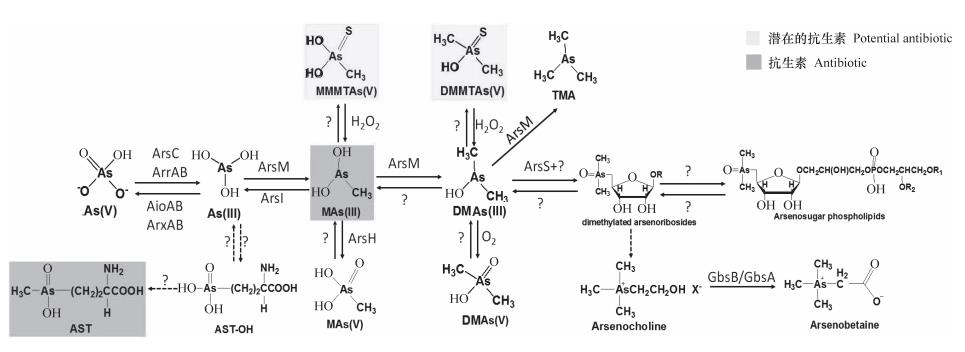
图1 土壤中砷的生物转化途径
1砷的生物氧化还原
微生物砷氧化过程加速自然环境中毒性和流 动性更强的As(III)向毒性和流动性更小且易吸 附在矿物表面的As(V)转化,降低土壤中砷的生 物有效性和毒性,因此微生物砷氧化被认为是降 低砷的生物有效性, 从而控制砷污染的重要途径 之一。微生物的砷还原将固定于矿物中的As(V) 释放并还原为移动性强的As(III),这一作用机 理可运用于尾矿等区域砷污染严重的土壤砷的去 除。对参与微生物砷的生物氧化还原过程的微生物 类型及其氧化还原机制研究得较清楚,已有多篇综 述对该过程进行了较为详细的介绍[6-7],本文不再 赘述。
土壤动物与微生物植物一起构成复杂的土壤 食物网,在陆地生态系统中扮演着重要的角色。 它们在凋落物分解、 土壤中元素循环、生物多样 性的维持、土壤微团聚体的形成和污染物的降解 等诸多土壤生态过程中发挥了关键作用。土壤动 物肠道内的代谢及其营养元素富集过程改变特定 元素的生物有效性。 多项研究在蚯蚓Lumbricus rubellust、Dendrodrilus rubidus等的组织中检测到 了As(III)、As(V)、甘油砷糖(Oxo-Gly)、磷酸砷糖(Oxo-PO4)、硫酸砷糖(Oxo-SO4)、 砷甜菜碱( AB )、 MAs ( V )、二甲基砷酸
[DMAs(V)]和三甲基砷氧化物(TMAO)等
(表1),但无法确定这些不同形态砷的来源。动 物体内生活着各种各样的微生物,其与宿主长期共 存、协同进化,对宿主的健康、免疫、养分获取和 运动等诸多方面发挥着不可替代的作用。Wang等
[8]用As(V)处理蚯蚓28 d后,发现土壤中仍以 As(V)为主,而蚯蚓肠道和组织中As(III)是 砷的主要存在形式,砷氧化还原和解毒外排基因在 所有基因中占主导,砷甲基化去甲基化相关的基因 含量很低,说明在厌氧蚯蚓肠道中微生物对砷转化 以氧化还原为主。无论蚯蚓肠道或组织中有机砷是 直接来自环境还是复杂有机砷进入肠道后肠道微生 物对其的降解或转化,蚯蚓等土壤动物的活动均将 严重影响土壤或土壤孔隙水中砷的迁移转化、形态 和生物有效性。
植物通过磷酸转运蛋白(如水稻的OsPTs[14]) 吸收环境中的As( V)。大多数植物的根细胞能 将进入其内的As(V)迅速还原成As(III),例 如水稻的OsACR2[15],A. thaliana的HAC1[16]、 Pte ri s vit t at a 的 PvACR2 [ 17 ] 等可能参与了 As
( V )的还原。植物体利用水甘油通道蛋白( aquaglyceroporins , AQPs ) [ 18 ] 吸收的 As
( III),一部分被富含巯基的多肽—植物络合素
( Phytochelatin, PC)络合并被液泡膜上的ABC 转动子转入到液泡内[ 19 ],另一部分则通过木质 部往地上部运输(如水稻的Lsi2[20]和A. thaliana 的肌醇转运蛋白 AtINT2 和 AtINT4 参与了该过 程[21])。
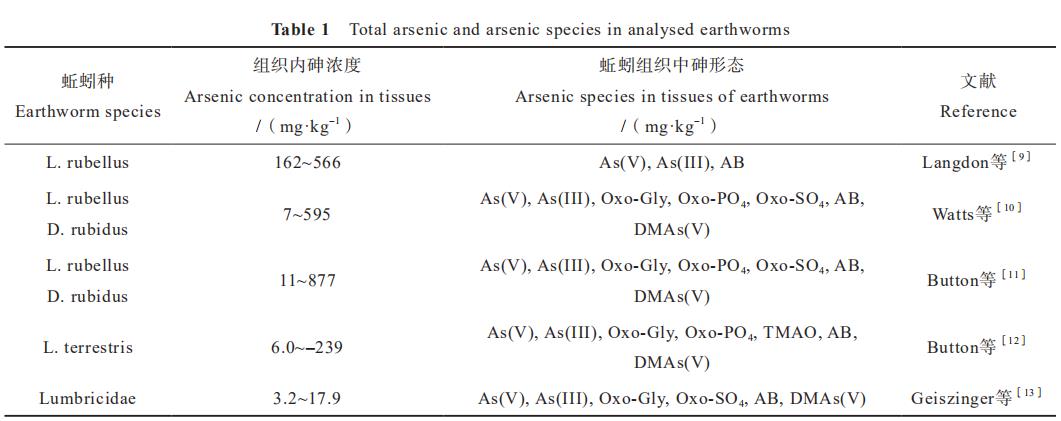
表1 已分析的蚯蚓中的砷总量及形态
2 砷的生物甲基化及去甲基化
砷甲基化可以将毒性较大的无机砷转化成毒性 更小的甲基化砷,虽然中间产物可能是毒性更大的 MAs( III),但该中间产物极不稳定,短时间内 即被氧化成MAs(V)[22]。对厌氧微生物而言, 砷甲基化过程多半产生挥发性甲基化砷从而降低环 境中砷浓度,砷甲基化可能是一个解毒过程[23]。
砷在生物体内甲基化过程是由砷甲基化酶催 化,以S- 腺苷甲硫氨酸( SAM)为甲基供体,巯 基在还原中起重要作用。 Marapakala等[ 24 ] 将真 核生物Cyanidioschyzon sp. 5508的CmArsM中三 个保守半胱氨酸依次突变后,蛋白荧光检测证明 As(GS)3和MAs(GS)2较As(III)有更快的结 合能力,这暗示着谷胱甘肽化砷可能是CmArs M 的首选底物,并通过分析与苯亚砷酸盐[ P hAs
(III)]或还原性洛克沙砷[Rox(III)]结合的 CmArsM晶体结构,推测CmArsM的砷甲基化过程 是一个二硫键形成和断裂的级联反应[25]。
土壤中存在具有砷挥发能力的细菌或真菌已被 证实[26],真核藻类也具有将无机砷转化为气态砷的能力[27]。土壤酸碱度、氧化还原电位、含水率 和温度等理化性质以及砷浓度、形态和组成等均直 接或间接影响着砷的挥发。但在自然条件下土壤砷 挥发量很低,可通过调节土壤环境或者通过基因工 程的手段改造土壤生物,提高砷挥发水平从而达到 土壤砷的生物修复[28-29]。
另一方面进入土壤的大部分有机砷将被降解成 毒性更强的无机砷。Yoshinaga等[30]从MAs(V) 除草剂处理的高尔夫球场土壤中分离到两株菌,一 株布克氏菌(Burkholderia)可将MAs(V)还原 成MAs(III),而另一株链霉菌(Streptomyces) 可将MAs(III)脱甲基成As(III),这两种菌中 任何一种单独存在均不能完成从MAs( V)到As
( III)的脱甲基过程,暗示土壤微生物群落催化 脱甲基化反应是由还原和去甲基化两个步骤组成。 针对此过程的第二步,Yoshinaga等得到一个MAs
( III)去甲基化酶( ArsI),该酶可以催化依赖 Fe2+的MAs(III)去甲基化反应,此外,ArsI还能 打断Rox(III)和其他含砷芳香族化合物的C-As键
[31],且在原核生物中广泛存在[32]。这表明有机 砷降解普遍存在于自然界中,暗示着去甲基化过程 在砷的土壤循环中发挥重要作用。
3 砷的其他有机化过程
除人工合成的有机砷外,土壤中还检测到了 砷糖、砷甜菜碱及砷胆碱等多种生物来源的有机 砷[33]。近期在复杂有机砷的生物合成机制方面究 取得了一系列进展。
自从 1981 年 Edmonds 和 Francesconi [ 34 ] 在褐藻( Ecklonia radiate)中第一次分离鉴定 到砷糖。迄今,人们已至少鉴定出了氧代砷 糖( Oxo-arsenosugars )、硫代砷糖( Thio- arse nosugars )、三甲基砷糖( t ri me thyla te d arsenosugars)三大类共20多种砷糖。砷糖可能是 砷糖磷脂合成的起点,砷胆碱和砷甜菜碱可能是砷 糖降解产物[1,35]。但因前期仅在基因组未测序且 遗传操作复杂的海洋生物中检测到砷糖,无法开展 砷糖生物合成分子机制的研究。Xue等[36-37]发现 纯培养的集胞藻和念珠藻均能将As(V)转化成砷 糖及结构更复杂的砷糖磷脂,并基于Zhu等[1,35] 对砷糖合成机制的推测,依次敲除集胞藻的arsM 和arsS两个基因,用不同形态砷处理突变体及野生 型,发现砷糖的合成是从DMA开始的,在砷糖合 成过程中ArsM的功能是提供DMAs,ArsS可能将 一个腺苷自由基转移给DMA同时将腺嘌呤切掉形 成另一个自由基。双表达arsM和arsS的大肠杆菌不 能将As(V)或As(III)转化成砷糖,暗示蓝细 菌中砷糖的合成还需要其他所特有基因[38-39]。总 之,砷糖是砷生物地球化学循环的重要组成部分, 它广泛存在于海洋和陆生生物中,但其存在的生物 学意义和生态学功能还有待进一步研究。
砷甜 菜碱是目前 公认的 毒性最小 的含砷化 合物,但其生物学功能仍不清楚。 Hof fmann 等[40]研究表明B. subtilis的甜菜碱合成酶(GbsB/ GbsA)同样能将砷胆碱转化成砷甜菜碱。此外, 环境中的Shewanella putrefaciens将MAs(V)和 DMAs( V)分别硫基化成毒性更强的单甲基单 硫砷酸[ MMMTAs ( V )]和二甲基单硫砷酸
[DMMTAs(V)][41]。
4从砷污染到抗生素抗性基因
4.1抗生素抗性基因及共选择抗性机制
抗生素的发现与使用拯救了千千万万的生命, 是人类医疗史上划时代的成就。但随着抗生素的过 度使用和滥用,ARGs的多样性与丰度在环境中不 断增加[ 42-43 ]。当多种ARGs同时转移到一种病原 菌使其带有多重抗性,则会形成"超级细菌",从 而使抗生素不再有效[44]。不断发展的采矿冶金、 化工和农业将越来越多的重金属释放到土壤,加上抗生素的滥用,加剧了环境中抗生素及重金属协同 污染的概率。重金属通过共选择的方式促进ARGs 在生态系统中的扩散,并且这种共选择具有普遍 性,不局限于几种重金属或者微生物[45]。多项野 外调查研究显示,土壤中的重金属与ARGs间存在 明显相关性,但不同金属对ARGs丰度和多样性的 影响存在差异[46-47]。现在研究认为共选择抗性机 制有四种(图2):
共抗性(co-resistance)是指ARGs或/和重金 属抗性基因位于同一质粒、转座子、整合子上或染 色体邻近位置。如Ghosh等[48]从马流产沙门氏菌 提到的质粒同时抗氨苄青霉素、砷、镉、铬和汞。 然而,必须指出的是,抗金属质粒早于人类使用抗 生素前已经出现。Mindlin等[49]认为含有ARGs和 汞抗性基因并携带转座子的整合子的出现是一个 相对较新的现象,因为冻土源假单胞菌尽管缺乏 ARGs盒但含有许多转座子与当今细菌中发现的关 系密切。此外,从抗生素时代前的肠杆菌科培养标 本中提取的R质粒中没有可转移的ARGs[50]。这表 明重金属与抗生素的协同进化一直在发生。现在需 要解决的问题是早已存在的金属抗性基因是在何时 如何获得ARGs的,这个时间点是否在抗生素广泛 使用前,金属或类金属增加ARGs丰度和多样性的 分子机制。对参与基因水平转移的可移动遗传元件 大尺度测序将有助于我们深刻理解环境中金属与抗 生素的共抗机制。
交叉抗性(cross-resistance)指微生物利用同 一套抗性系统同时对抗生素和重金属产生抗性, 当不同的抗菌剂胁迫同一种细菌时,细菌对一种 抗菌剂产生耐药性的同时对另一种抗菌剂也产生 抗性,即细菌使用相同机制排出结构不同的化合 物。例如,Listeria monocytogenes的多药物抗性
(multiple-drug resistance,MDR)泵可同时外排 抗生素和重金属[ 51 ]。编码MDR泵的基因广泛存 在,但目前还没有研究能阐明这种交叉抗性的分子 机制:MDR泵基因的进化、MDR泵基因的表达是 否受多种抗生素或金属的调控、MDR泵是如何同 时识别并排出结构差异较大的抗菌剂。
共调控(co-regulation)指一系列能对金属或 抗生素暴露做出反应的转录和翻译应答系统关联在 一起形成协同反应, 如绿脓单胞菌的双组份传感 器蛋白CzcS负责该菌对锌和亚胺培南的抗性[52]。
在大肠杆菌Bae RS调控八个基因的表达: mdt A, mdtB, mdtC, mdtD, baeS, baeR, acrD和spy, 其中 MdtABC 与外膜孔蛋白 TolC 形成一个 RND
(resistance, nodulation and division)型跨膜外排 蛋白复合体,AcrD是AcrB的同源蛋白形成RND型 转运复合体的内膜部分,Spy是一种外周质蛋白在 体外表现出不依赖于ATP的伴侣活性,MdtD的功 能未知[53]。Hao等[54]发现识别Cu(II)的MarR 降低了大肠杆菌多抗生素抗性调控操纵子marRAB 的表达从而导致了大肠杆菌对多种抗生素的抗性。 经过砷、铜和锌的诱导,Enterobacter LSJC7表现 出对四环素的抗性,同时LSJC7 也含有baeRS, acrD,mdtABC,soxS,acrAB,tolC,marRAB 等基因,可能像在大肠杆菌中一样锌和铜分别通 过调控BaeRS和MarR增加了LSJC7对四环素的抗 性,但砷如何增强LSJC7对四环素抗性的机制有待 进一步探讨[55]。
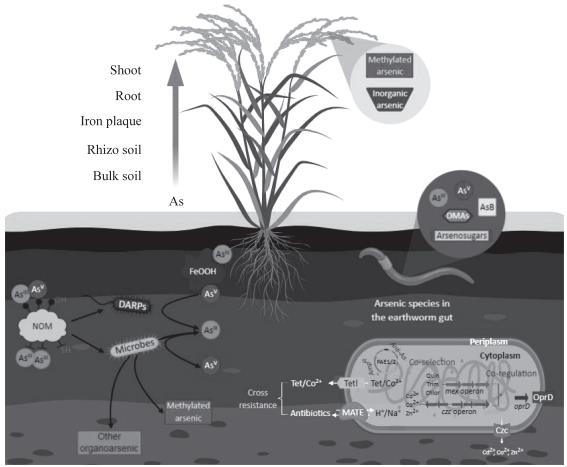
NOM,天然有机物(natural organic matter);DARP,异化砷呼吸原核生物(Dissimilatory As(V)-respiring prokaryotes); Tetl,四环素泵出蛋白(tetracycline efflux protein);MATE,多药物和有毒化合物外排蛋白(multidrug and toxic compound extrusion);PAE1/2,马流产沙门氏菌157/158抗性质粒(Salmonella abortus equi 157/158 resistance plasmid)[48];Quin,喹 诺酮(quinolone);Tet,四环素(tetracycline);Trim,甲氧苄氨嘧啶(trimethoprim);Chlor,氯霉素(chloramphenicol); AmpR,氨苄青霉素抗性基因(Ampicillin resistance gene);Anti-As,砷抗性基因
图2 土壤微生物和动物对砷形态的转化及砷与抗生素抗性关联可能的分子机制
生物膜感应(Biofilm induction)指重金属或 抗生素的加入刺激微生物形成一种包裹于细胞外 的多聚物, 增强了其对重金属和抗生素的抗性。 金属和抗生素在生物膜基质中的滞留和少量存留 细胞(persister cells)的存在可能是浮游生物细 胞和生物膜对高浓度抗菌剂产生时间依赖耐受性 的原因[ 56 ]。向浮游生物细胞中添加金属或抗生 素均可刺激胞外聚合物(extracellular polymeric substances)的产生,从而导致细胞粘附,最终形 成生物膜[57-58]。此外生物膜内被保护的环境和微 生物间的近距离分布也为基因的横向转移提供了 便利。
Yamamura等[ 59 ]研究抗生素对砷氧化还原和 砷氧化菌多样性的影响,发现有氧As(V)还原不 受抗生素影响,但有氧As(III)氧化菌对某些特 定的抗生素(如氯霉素)敏感,这表明大多数抗砷 菌可能同时抗抗生素。Wang等用As(V)和磺胺甲恶唑处理蚯蚓,发现砷明显增加了蚯蚓肠道微生 物中特定ARGs和移动遗传元件的出现频率(数据 未发表),推测进入蚯蚓肠道的As(V)被其微生 物转化成毒性更强的As(III)后富集了双抗(抗 砷和抗生素)的细菌,但具体机制仍需研究。总 之,在土壤生态系统中土壤动物及微生物面临多重 胁迫,土壤中砷的生物转化(尤其是转化成毒性更 强的砷化合物)通过干扰微生物群落结构导致了 ARGs丰度和多样性的改变,不同形态砷和抗生素 抗性的耦合关系及机制需要进一步研究。
4.2 含砷抗生素
近期Li等[60]提出有机砷可能是最原始的抗生 素,根据Waksman[ 61 ] 对抗生素的定义,一种微 生物产生的化学物质能抑制细菌和其他微生物的 生长甚至杀死它们, MAs( III)可作为一种抗生 素,低浓度MAs( III)即可抑制大肠杆菌的生长
[ 62 ]。土壤微生物群落中有些携带arsM的细菌产 生剧毒的MAs(III)[22]。此外已发现有四种菌P. putida KT2440[30]、Burkholderia sp. MR1[30]、 Shewanella putrefaciens 200 和Sinorhizobium meliloti RM1021[62]通过将MAs(V)还原成MAs
(III)获得生存竞争优势。而为了在MAs( III) 选择 压力 下生 存,其 他土 壤细 菌进化 出抗 MAs
( III)的基因, Bacillius MD1 利用ArsI将MAs
(III)去甲基化成As(III)解毒[31],P. putida用 ArsH和NADP+将MAs(III)氧化成毒性小的MAs
(V)[62],Campylobacter jejuni编码ArsP [MAs
(III)排出透性酶]将细胞内的毒性强的三价有机 砷排出[63]。
砷代草丁膦( AST )是水稻根际微生物 Burkholderia gladioli GSRB05产生的新型砷有机 物[64]。Nadar等[65]发现AST抑制谷胱酰胺合成酶 的活性,是一种含砷的广谱抗生素,能同时有效对 抗革兰氏阳性菌和革兰氏阴性菌,其毒性与MAs
(III)相当,较As(III)和草丁膦的毒性更强。P. putida KT2440的ArsN可将AST的α-氨基乙酰化阻 止AST与谷胱酰胺合成酶的结合。AST具备治疗人 类结核病的潜力,但该抗生素在动物和人类中的有 效性和毒性还需要进一步测试[66]。总之环境中总 有一些微生物产生含砷抗生素抑制或杀死其他微生 物,而也有一部分微生物进化出特定的机制来适应 这种环境。
4.3未来研究的方向
土壤是一个极其复杂的介质,土壤理化性质影 响土壤动物和微生物种类及其代谢过程,土壤中的 植物、动物和微生物可以吸收转化或代谢砷。就土 壤砷转化过程而言,无机砷一直是砷代谢的终点, 无论是人工合成还是自然产生的复杂或简单有机 砷,在自然条件下大多会被慢慢降解,最终产物是 无机砷;从微生物转录调控来看,参与砷氧化还原 过程的基因大多是受砷调控的,参与甲基化和其他 有机化过程的基因多是组成性表达的,暗示环境中 砷的氧化还原过程是砷代谢的主流。然而随着精确 分析化学和现代分子生物学技术的进步,越来越多 新型含砷化合物以及合成或降解这些化合物的生物 或基因被发现。因这些含砷化合物在土壤中的含量 较低,我们还无法知道生物为什么消耗能量产生这 些复杂的有机砷、它们存在的生物学意义或生态学 功能以及在食物链上的传递规律,这一系列问题有 待进一步探讨。在分子水平上分析不同形态砷与土 壤各组分之间的相互作用机制,土壤中微生物的种 类及含量对砷化学行为的影响等有助于我们更好地 理解土壤砷转化过程,为土壤砷污染修复提供重要 的理论依据。此外在探明土壤微生物对砷转化分子 机制的基础上进一步在群落水平上阐明其微生物生 态学机制,为发展定向调控土壤中功能微生物的技 术途径奠定基础。总之这些研究成果可被用来有效 控制土壤—植物系统中砷的转化和积累,最终达到 降低土壤砷污染风险的目标。
很明显大量抗菌药物的消耗与抗生素抗性基 因的数量及种类增加显著相关,一系列其他污染物 也驱动ARGs的选择。考虑到环境中普遍存在砷和 抗生素等复合污染,微生物为抵抗砷和抗生素双重 污染势必要进化出一套同时抗砷和抗生素的抗性系 统,这些抗性菌株及其携带的ARGs更易在环境中 传播扩散, 将威胁人类健康和生态安全。 然而现 在的研究未能充分解决在生态相关框架内(如当 定量ARGs在特定环境的丰度、转移和保留时区分 暴露于砷的微生物种群的群落结构改变) 砷暴露 是增加ARGs传播和出现频率的一种重要机制。微 生物基因组学、生理学和生物化学的最新进展可 为精确测定砷-ARGs相互作用提供依据。此外,很 少有研究将共选择的分子机制与种群和群落水平上 的效应联系起来,因此有必要深入研究复合污染对环境ARGs的富集和传播机制以全面评估砷污染在 维持和传播环境中ARGs的作用。并对砷和抗生素 之间复杂的相互关系进行详细描述,以便系统理解 ARGs的持续性和增殖。同时,一些新型的砷生物 转化产物具有抗生素性能,对含砷抗生素的生物合 成、毒理学及降解机制的研究无疑也是一个新的研 究方向。
参 考 文 献
[ 1 ] Zhu Y G,Yoshinaga M,Zhao F J,et al. Earth abides arsenic biotransformations. Annual Review of Earth and Planetary Sciences,2014,42(1):443—467
[ 2 ] Chen Z,Zhu Y G,Liu W J,et al. Direct evidence showing the effect of root surface iron plaque on arsenite and aresnate uptake in rice(Oryza sativa) roots. New Phytologist,2005,165(1):91—97
[ 3 ] Sun G X,Williams P N,Carey A M,et al. Inorganic arsenic in rice bran and its products are an order of
magnitude higher than in bulk grain. Environmental Science & Technology,2008 , 42 ( 19 ): 7542 — 7546
[ 4 ] Wang H T,Zhu D,Li G,et al. Effects of arsenic on gut microbiota and its biotransformation genes in earthworm Metaphire sieboldi. Environmental Science
& Technology,2019,53:3841—3849
[ 5 ] Jia Y,Huang H,Chen Z,et al. Arsenic uptake by rice is influenced by microbe-mediated arsenic redox changes in the rhizosphere. Environmental Science & Technology,2014,48(2):1001—1007
[ 6 ] 杨婧,朱永官. 微生物砷代谢机制的研究进展. 生态毒 理学报,2009,4(6):761—769
Yang J,Zhu Y G. Progress in study of mechanisms of microbial arsenic transformation in environment
(In Chinese). Journal of Ecotoxicology,2009,4
(6):761—769
[ 7 ] 蔡林,王革娇. 抗砷性微生物及其抗砷分子机制研究 进展. 微生物学通报,2009,36(8):1253—1259
Cai L, Wang G J. Advance on studies of arsenic- resistant microorganisms and molecular mechanisms
(In Chinese). Microbiology Bulletin, 2009: 36
(8):1253—1259
[ 8 ] Wang X,Chen X,Yang J,et al. Effect of microbial mediated iron plaque reduction on arsenic mobility in paddy soil. Journal of Environmental Sciences, 2009,21(11):1562—1568
[ 9 ] Langdon C J, Piearce T G, Feldmann J, et al. Arsenic speciation in the earthworms Lumbricus rubellus and Dendrodrilus rubidus. Environmental Toxicology and Chemistry,2003,22(6):1302—
1308
[10] Watts M J,Button M,Brewer T S,et al. Quantitative arsenic speciation in two species of earthworms from a former mine site. Journal of Environmental Monitoring,2008,10(6):753—759
[11] Button M , Jenkin G R T , Harrington C F , et al. Arsenic biotransformation in earthworms from contaminated soils. Journal of Environmental Monitoring,2009,11(8):1484—1491
[12] Button M,Moriarty M M,Watts M J,et al. Arsenic speciation in field-collected and laboratory-exposed earthworms Lumbricus terrestris. Chemosphere, 2011,85(8):1277—1283
[13] Geiszinger A , Goessler W , Kuehnelt D. Determination of arsenic compounds in earthworms. Environmental Science & Technology, 1998 , 32
(15):2238—2243
[14] Kamiya T,Islam M R,Duan G,et al. Phosphate deficiency signaling pathway is a target of arsenate and phosphate transporter OsPT1 is involved in As accumulation in shoots of rice. Soil Science and Plant Nutrition,2013,59(4):580—590
[15] Duan G L, Zhou Y, Tong Y P, et al. A CDC25 homologue from rice functions as an arsenate reductase. New Phytologist,2007,174(2):311— 321
[16] Chao D Y,Chen Y,Chen J,et al. Genome-wide
association mapping identifies a new arsenate reductase enzyme critical for limiting arsenic accumulation in plants. PLoS Biology,2014,12(12):e1002009.
DOI: 10. 1371/journal. pbio. 1002009
[17] D u a n G L , Z h u Y G , T o n g Y P , e t a l . Characterization of arsenate reductase in the extract of roots and fronds of Chinese brake fern,an arsenic hyperaccumulator. Plant Physiology, 2005 , 138
(1):461—469
[18] Liu Z,Shen J,Carbrey J M,et al. Arsenite transport by mammalian aquaglyceroporins AQP7 and AQP9. Proceedings of the National Academy of Sciences, 2002,99(9):6053—6058
[19] Song W Y,Yamaki T,Yamaji N,et al. A rice ABC transporter,OsABCC1,reduces arsenic accumulation in the grain. Proceedings of the National Academy of Sciences,2014,111(44):15699—15704
[20] Ma J F,Yamaji N,Mitani N,et al. Transporters of arsenite in rice and their role in arsenic accumulation in rice grain. Proceedings of the National Academy of
Sciences,2008,105(29):9931—9935
[21] Duan G L, Hu Y, Schneider S, e t a l . Inositol transporters AtINT2 and AtINT4 regulate arsenic accumulation in Arabidopsis seeds. Nature Plants, 2015,2(1):15202
[22] Qin J , Rosen B P , Zhang Y , et al. A rsenic detoxification and evolution of trimethylarsine gas by a microbial arsenite S-adenosylmethionine methyltransferase. Proceedings of the National
Academy of Sciences,2006,103(7):2075—2080
[23] Wang P,Sun G,Jia Y,et al. A review on completing arsenic biogeochemical cycle:Microbial volatilization of arsines in environment. Journal of Environmental Sciences,2014,26:371—381
[24] Marapakala K, Qin J, Rosen B P. Identification o f c a t a l y t i c r e s i d u e s i n t h e A s ( I I I ) S-adenosylmethionine methyltransferase. Biochemistry 2012,51(5):944—951
[25] Marapakala K, Packianathan C, Ajees A A, e t al. A disulfide-bond cascade mechanism for arsenic
( III) S-adenosylmethionine methyltransferase.
Acta Crystallographica Section D : Biological Crystallography,2015,71(Ⅲ):505—515
[26] Jia Y, Huang H, Sun G X, et al. Pathways and relative contributions to arsenic volatilization from rice plants and paddy soil. Environmental Science &
Technology,2012,46(15):8090—8096
[27] Guo Y,Xue X,Yan Y,et al. Arsenic methylation by an arsenite S-adenosylmethionine methyltransferase from Spirulina platensis. Journal of Environmental Science,2016,49:162-168
[28] Meng X Y , Qin J , Wang L H , et al. Arsenic biotransformation and volatilization in transgenic rice. New Phytologist,2011,191:46—59
[29] Zhu Y G , Rosen B P. Perspectives for genetic engineering for the phytoremediation of arsenic- contaminated environments: from imagination to reality? Current Opinion in Biotechnology,2009,20
(2):220—224
[30] Yoshinaga M, Cai Y, Rosen B P. Demethylation of methylarsonic acid by a microbial community. Environmental Microbiology, 2011 , 13 ( 5 ): 1205—1215
[31] Yoshinaga M , Rosen B P. A C · As lyase for degradation of environmental organoarsenical herbicides and animal husbandry growth promoters.
Proceedings of the National Academy of Sciences,
2014,111(21):7701—7706
[32] Yan Y,Ye J,Xue X M,et al. Arsenic demethylation by a C · As lyase in cyanobacterium Nostoc sp. PCC 7120. Environmental Science & Technology,2015, 49:14350—14358
[33] Huang J H, Hu K N, Decker B. Organic arsenic in the soil environment:Speciation,occurrence, transformation, and adsorption behavior. Water, Air,and Soil Pollution,2011,219(1/4):401—
415
[34] Edmonds J S,Francesconi K A. Arseno-sugars from brown kelp(Ecklonia radiata)as intermediates in cycling of arsenic in a marine ecosystem. Nature, 1981,289(5798):602—604
[35] Zhu Y, Xue X, Kappler A, et al. Linking genes to microbial biogeochemical cycling:Lessons from arsenic. Environmental Science & Technology, 2017,51:7326—7339
[36] Xue X M,Raber G,Foster S,et al. Biosynthesis of arsenolipids by the cyanobacterium Synechocystis sp. PCC 6803. Environmental Chemistry,2014,11
(5):506—513
[37] Xue X M , Yan Y , Xiong C , et al. Arsenic biotransformation by a cyanobacterium Nostoc sp. PCC 7120. Environmental Pollution,2017,228:111—
117
[38] Xue X M , Ye J , Raber G , et al. Arsenic methyltransferase is involved in arsenosugar biosynthesis by providing DMA. Environmental Science & Technology,2017,51(3):1224—1230
[39] Xue X M,Ye J,Raber G,et al. Identification of steps in the pathway of arsenosugar biosynthesis. Environmental Science & Technology, 2019 , 53
(2):634—641
[40] Hoffmann T, Warmbold B, Smits S H J, et al. Arsenobetaine: an ecophysiologically important organoarsenical confers cytoprotection against osmotic stress and growth temperature extremes. Environmental Microbiology,2018,20(1):305—323
[41] C h e n J , R o s e n B P . O r g a n o a r s e n i c a l Biotransformations by Shewanella putrefaciens. Environmental Science & Technology,2016,50: 7956—7963
[42] 朱永官,陈青林,苏建强,等. 环境中抗生素与抗性 基因组的研究. 科学观察,2017,12(6):60—72
Zhu Y G,Chen Q L, Su J Q, et al. The study of antibiotics and resistance genomes in environment(In Chinese). Science Focus,2017,12(6):60—72
[43] 朱永官,欧阳纬莹,吴楠,等. 抗生素耐药性的来源 与控制对策. 中国科学院院刊,2015,30(4):509- 516
Zhu Y G, Ouyang W Y, Wu N, et al. Antibiotic resistance:Sources and mitigation(In Chinese). Bulletin of Chinese Academy of Sciences,2015,30
(4):509-516
[44] Forsberg K J,Reyes A,Wang B,et al. The shared antibiotic resistome of soil bacteria and human pathogens. Science, 2014 , 337 ( 6098 ): 1107 —
1111
[45] Seiler C,Berendonk T U. Heavy metal driven co- selection of antibiotic resistance in soil and water bodies impacted by agriculture and aquaculture. Frontiers in Microbiology,2012,3:399. DOI: 10. 3389/fmicb. 2012. 00399
[46] Zhu Y, Johnson T A, Su J, et al. Diverse and abundant antibiotic resistance genes in Chinese swine farms. Proceedings of the National Academy of Sciences,2013,110(9):3435—3440
[47] Zhu Y,Gillings M,Simonet P,et al. Microbial mass movements. Science, 2017 , 357 ( 6356 ): 1099- 1100
[48] Ghosh A , Singh A , Ramteke P W , et al. Characterization of lar ge plasmids encoding resistance to toxic heavy metals in Salmonella abortus equi. Biochemical and Biophysical Research
Communications,2000,272:6—11
[49] Mindlin S,Minakhin L,Petrova M,et al. Present- day mercury resistance transposons are common in bacteria preserved in permafrost grounds since the Upper Pleistocene. Research in Microbiology,2005, 156:994—1004
[50] Hughes V M, Datta N. Conjugative plasmids in bacteria of the 'pre-antibiotic'era. Nature,1983, 302(5910):725
[51] Mata M T,Baquero F,Pérez-Díaz J C. A multidrug efflux transporter in Listeria monocytogenes. FEMS Microbiology Letters,2000,187(2):185—188
[52] Perron K,Caille O,Rossier C,et al. CzcR-CzcS, a two-component system involved in heavy metal and carbapenem resistance in Pseudomonas aeruginosa.
Journal of Biological Chemistry,2004,279(10):
8761—8768
[53] Wang D , Fierke C A. The BaeSR regulon is involved in defense against zinc toxicity in E. coli. Metallomics,2013,5(4):372—383
[54] Hao Z,Lou H,Zhu R,et al. The multiple antibiotic resistance regulator MarR is a copper sensor in Escherichia coli. Nature Chemical Biology,2014, 10:21—28
[55] Chen S,Li X,Sun G,et al. Heavy metal inducedantibiotic resistance in bacterium LSJC7. International Journal of Molecular Sciences,2015,16(10): 23390—23404
[56] Harrison J J,Turner R J,Ceri H. Persister cells, the biofilm matrix and tolerance to metal cations in biofilm and planktonic Pseudomonas aeruginosa.
Environmental Microbiology,2005,7(7):981—
994
[57] García-Meza J V,Barrangue C,Admiraal W. Biofilm formation by algae as a mechanism for surviving on mine tailings. Environmental Toxicology and Chemistry,2005,24(3):573—581
[58] Wuertz S,Spaeth R,Hinderberger A,et al. A new method for extraction of extracellular polymeric substances form biofilms and activated sludge suitable for direct quantification of sorbed metals. Water Science and Technology, 2001 , 43 ( 6 ): 25 — 31
[59] Yamamura S,Watanabe K,Suda W,et al. Effect of antibiotics on redox transformations of arsenic and diversity of arsenite-oxidizing bacteria in sediment microbial communities. Environmental Science & Technology,2014,48(1):350—357
[60] Li J,Pawitwar S S,Rosen B P. The organoarsenical biocycle and the primordial antibiotic methylarsenite. Metallomics,2016,8:1047—1055
[61] Waksman S A. What is an antibiotic or an antibiotic substance? Mycologia,1947,39(5):565—569
[62] Chen J, Bhattacharjee H, Rosen B P. ArsH is an organoarsenical oxidase that confers resistance to trivalent forms of the herbicide monosodium methylarsenate and the poultry growth promoter
ro xa rsone . Mol e c ul a r Mi c rob i ol og y , 20 15 , 96
(5):1042—1052
[63] Chen J, Yoshinaga M, Rosen B P. The antibiotic action of methylarsenite is an emergent property of microbial communities. Molecular Microbiology, 2019,111(2):487—494
[64] Iuramata M,Sakakibara A F,Kataoka D R,et al. Arsinothricin ,a novel organoarsenic species produced by a rice rhizosphere bacterium. Environmental Chemistry,2016,13(4):723—731
[65] Na da r V S , Ch e n J , Dhe e m a n D S , e t a l . Arsinothricin , an arsenic-containing non- proteinogenic amino acid analog of glutamate,is a broad-spectrum antibiotic. Communications Biology, 2019 , 2 ( 1 ): 131. https://doi. org/10. 1038/
s42003-019-0365-y
薛喜枚1 朱永官1,2
(1 中国科学院城市环境研究所,中国科学院城市环境与健康重点实验室,厦门 361021)
(2 中国科学院生态环境研究所中心,城市与区域生态国家重点实验室,北京 100085)
来源:土壤学报

 京公网安备 11011102002334号
京公网安备 11011102002334号